肝素是临床最重要的抗凝药物之一,目前主要从猪小肠中提取,存在供应链不稳定、批次异质性和病原污染风险。,针对肝素生物合成中的关键限速酶——鸡源6-O-磺酸基转移酶(Ga6OST1),首先在毕赤酵母中通过分泌工程实现了高效表达,再结合半理性设计获得三突变体A77R/V107L/V248T。该变体的比活提高2.4倍,催化周转数(kcat)提升2.3倍,37°C半衰期延长5.3倍,熔融温度提高2.77°C。分子动力学模拟揭示,突变通过引入盐桥、强化疏水堆积和氢键网络,增强了酶的结构刚性。最终,该突变体对N,2-O-硫酸化肝素前体的6-O-硫酸化程度达到75.7%,接近商业猪肠肝素水平。这项研究为生物工程化肝素的低成本、大规模生产提供了关键酶模块。

肝素:动物提取的“金标准”背后的隐患
肝素是一种高度硫酸化的线性糖胺聚糖,近百年来一直是临床抗凝治疗的基石。但它目前主要从猪小肠黏膜中提取,不仅依赖养殖业,还存在供应链波动、批次间结构不均一以及潜在的病原污染(如非洲猪瘟、朊病毒)等风险。化学合成肝素因结构复杂、成本极高而不可行。因此,利用酶法体外合成“生物工程肝素”成为理想替代方案。
肝素的生物合成需要一系列磺酸基转移酶依次修饰,其中6-O-磺酸基转移酶(6OST) 催化将硫酸基从PAPS转移到葡萄糖胺6位羟基上。这个修饰直接影响肝素的抗凝血活性——它与3-O-硫酸化共同构成抗凝血酶结合的五糖关键结构。然而,6OST1(尤其是来自鸡的Ga6OST1)的催化效率低、热稳定性差,成为整个合成路径的限速步骤。以往研究中,人们甚至需要同时使用6OST1和6OST3两个酶才能达到足够的硫酸化程度,大大增加了工艺复杂性。
首先,为什么选择毕赤酵母而不是大肠杆菌?
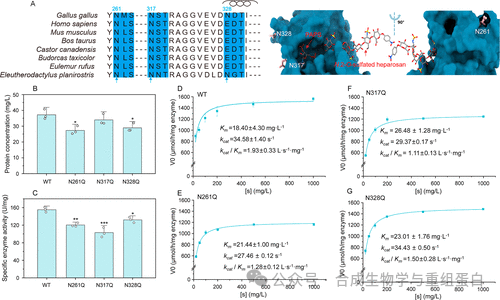
作者比较了MBP标签融合的Ga6OST1在毕赤酵母和大肠杆菌中的表达。结果显示,毕赤酵母来源的酶比活力(155.0 U/mg)略高于大肠杆菌来源(132.1 U/mg)。进一步的序列分析发现,Ga6OST1存在三个潜在的N-糖基化位点(N261、N317、N328)(图2A)。通过逐一突变为谷氨酰胺来消除糖基化,结果发现N261Q和N328Q突变导致蛋白分泌量下降约23-27%,表明这两个位点的糖链可能有助于折叠或分泌稳定性。而N317Q突变虽然蛋白量未明显下降,但比活力降低了33.6%,且对底物的Km值增加了43.9%(图2B-G),暗示该位点的糖基化可能邻近PAPS结合口袋,影响催化效率。尽管如此,所有糖基化突变体的残余活性仍高于大肠杆菌表达的野生型。因此,团队选择毕赤酵母作为表达宿主,并保留天然糖基化位点。
模块化分泌工程:让酶“出得去、活得好”
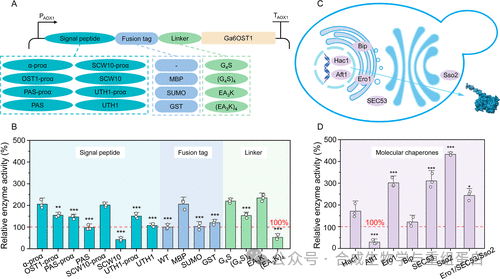
为了进一步提高胞外酶活,作者对分泌路径进行了系统优化(图3A)。首先比较了多种信号肽——包括酿酒酵母α因子、OST1信号肽、毕赤酵母内源PAS、SCW10、UTH1,以及将α因子的前导序列与后三者融合形成的嵌合体。结果显示,野生型α因子信号肽效果最佳,胞外活性达5.7 U/mL。随后筛选融合标签,发现MBP(麦芽糖结合蛋白)效果最优,远优于SUMO和GST。接着优化MBP与Ga6OST1之间的连接肽,发现短刚性连接肽(EA₃K)₁在稳定性和活性上最佳,胞外活性提升至6.6 U/mL。
接下来,团队共表达了多种分子伴侣:转录调节因子Aft1、Hac1,折叠因子BiP、Ero1,以及分泌运输辅助因子SEC53、Sso2(图3C)。其中Sso2(SNARE复合体组分,参与胞吐后期)效果最突出,将胞外活性进一步推高至12.0 U/mL,比未优化时提高约4.2倍(相对于无标签对照约2.1倍)(图3D)。同时共表达Ero1和SEC53也有一定提升,但组合共表达反而没有叠加效应,可能增加了代谢负担。
半理性设计:20个热点中筛出稳定突变
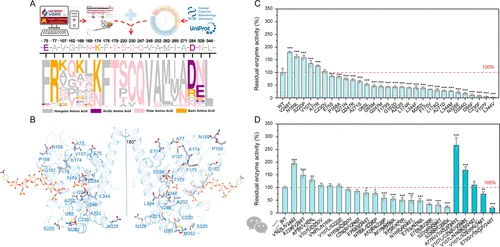
尽管分泌优化提升了产量,但Ga6OST1本身的热稳定性仍不足:37°C孵育2小时后残余活性仅约75%。为了提高酶自身的稳定性,团队采用半理性设计策略。首先用AlphaFold2预测Ga6OST1结构,结合HotSpot Wizard分析柔性残基和空腔,再对90条同源序列进行多序列比对和祖先序列重构,筛选出20个位于催化核心之外但可能影响局部堆积或静电网络的候选位点(图4A-B)。这些位点分布在酶的多个区域(图4B)。
将这些位点分别突变为更有利于疏水堆积或静电相互作用的氨基酸(如Val→Leu, Ala→Arg, Ser→Pro等),构建单点突变体库。在37°C热处理2小时后测定残余活性,发现多个突变体的热稳定性显著优于野生型(图4C)。其中V248T、N169L、S220P、V107L、A77R分别提高了1.8、1.6、1.5、1.3、1.2倍。将这些正向突变组合后,三突变体A77R/V107L/V248T表现出最佳的残余活性,比野生型提高了2.7倍(图4D)。该组合被命名为工程化MBP-Ga6OST1,用于后续详细表征。
酶学性质:比活、kcat、半衰期全面跃升
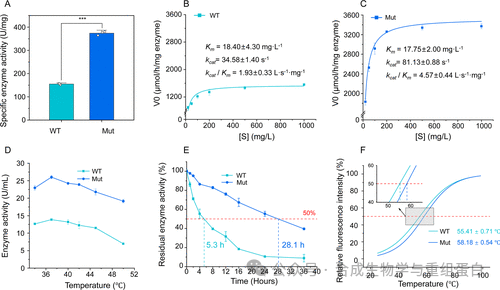
纯化后的工程酶与野生型进行对比(图5)。比活力从155.0 U/mg提升至374.1 U/mg(提高2.4倍,图5A)。动力学分析显示,工程酶的kcat从34.58 s⁻¹升至81.13 s⁻¹(提升2.3倍),而Km几乎没有变化(18.40 vs 17.75 mg/L),因此催化效率kcat/Km提升了2.4倍(图5B-C)。这表明突变主要提高了催化速率而非底物亲和力。
最适温度分析显示两者均在37°C最佳(图5D)。在37°C下的稳定性测试中,野生型半衰期仅为5.3小时,而工程酶达到28.1小时,延长了5.3倍(图5E)。差示扫描量热(thermal shift assay)显示工程酶的熔融温度Tm从55.41°C提高到58.18°C,增加了2.77°C(图5F),直接证明热稳定性的提升。
分子机制:盐桥、疏水堆积、氢键协同增强刚性
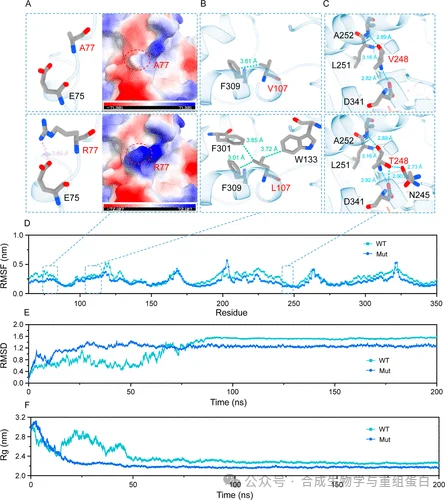
为了解释突变如何稳定酶结构,作者进行了200 ns的分子动力学模拟(图6)。A77R突变引入了一个长链带正电的精氨酸,与相邻的Glu75形成新的盐桥,增强局部静电网络(图6A)。V107L突变为体积更大的亮氨酸,埋入疏水核心,与Phe309、Trp133、Phe301等形成更强的疏水堆积,溶剂可及表面积(SASA)从11.475 Å⟡降至10.404 Å⟡,说明埋藏更深(图6B)。V248T突变为苏氨酸,引入两个额外的氢键,稳定了邻近α-螺旋及其与环区的相互作用(图6C)。
RMSF分析显示,工程酶在突变位点附近的残基柔性明显降低(图6D)。RMSD轨迹表明工程酶更快达到平衡,且整体波动更小(图6E)。回转半径Rg显示工程酶结构更紧凑、更早稳定(图6F)。这些模拟结果一致说明,三点突变协同作用增强了酶的整体刚性,从而提高了热稳定性和催化耐久性。
底物适应性:从非硫酸化到高度硫酸化,效率全面提升
6OST的一个独特优势是底物谱较宽,可以作用于不同硫酸化程度的肝素前体。作者测试了三种底物:非硫酸化的肝素糖(heparosan)、N-硫酸化肝素糖、N,2-O-硫酸化肝素糖(图7)。对于非硫酸化底物,野生型仅能将7.05%转化为6-O-硫酸化产物,而工程酶达到30.08%,效率提高4.26倍。对于N-硫酸化底物,工程酶转化率79.02%(野生型24.96%)。对于最接近最终产品的N,2-O-硫酸化底物,工程酶达到了75.70%的6-O-硫酸化程度,而野生型仅38.67%。值得注意的是,75.7%的6-O-硫酸化水平与商业猪肠肝素的约77%十分接近,表明工程酶具备了生产高质量生物工程肝素的潜力。
总结:突破肝素生物合成的“限速关”
本研究系统解决了Ga6OST1表达差、活性低、稳定性差三大难题:
表达优化:在毕赤酵母中利用α信号肽、MBP标签、短刚性连接肽,以及共表达Sso2分子伴侣,使胞外酶活提高约4.2倍。
半理性设计:基于进化分析和结构预测,筛选出20个热点,获得三突变体A77R/V107L/V248T,比活提高2.4倍,半衰期延长5.3倍,Tm提高2.77°C。
机制阐明:MD模拟揭示盐桥、疏水堆积、氢键网络协同增强结构刚性。
应用验证:工程酶对N,2-O-硫酸化底物的6-O-硫酸化程度达75.7%,接近天然肝素标准。
该工作不仅为肝素的绿色生物制造提供了一个高效、稳定的催化模块,也展示了“真核表达优化+半理性设计”改造磺酸基转移酶的通用策略。未来,将工程Ga6OST1与其它修饰酶(NDST、2OST、3OST)级联,有望实现全酶法合成结构均一的生物工程肝素,彻底摆脱对动物来源的依赖。
原文信息:Xinjing Wang, Yaxin Yu, Ping Dong, Daoan Wang, Ruirui Xu, Guocheng Du, Xintong Xi, Zhen Kang. ACS Synthetic Biology, 2026, 15, 2160-2172. DOI: 10.1021/acssynbio.6c00237
通讯作者:Zhen Kang(康振)、Xintong Xi(席心彤),江南大学
第一单位:江南大学糖化学与生物技术教育部重点实验室


 微信服务号
微信服务号
 微信订阅号
微信订阅号
 微信订阅号
微信订阅号